
表型可塑性
表型可塑性指生物體之行為、形態、生理為因應獨特環境所發生之表現型差異[1],這些差異涵蓋了環境引起的所有變化類型,如形態、生理、行為、物候等多方面的變化;這些變化在生物個體壽命中是永久性的,但也可能是暫時性的。這一術語最初是用來描述發育對於形態特性的影響,但現在更廣泛地用來描述對環境變化產生的如馴化(適應)、學習等的表型差異。[2]表型可塑性中環境差異誘導產生非連續表型的現象,也被稱為非遺傳多型性[3]。

一般來說,表型可塑性對於如植物等的較為靜態的生物更為重要。而對於大多數動物等可移動的生物而言,它們可以通過遷移遠離不利環境[4]。儘管如此,移動的生物也至少在某些方面具有一定程度的表型可塑性。豌豆蚜(學名:Acyrthosiphon pisum)即體現出顯著的表型可塑性:豌豆蚜的生殖方式可以在無性和有性生殖之間不斷轉換,且豌豆蚜在群落過於密集的時候長出翅膀。[5]
植物中的例子
[編輯]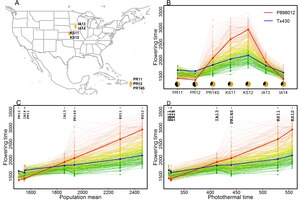
植物的表型可塑性,既涵蓋從生長階段過渡到生殖階段的時間調控,也包括植物應對不同營養程度的土壤的對根系不同的養料分配,還囊括環境對於植物個株的種子的大小的影響[7],環境對於葉子形狀、大小和厚度的改變等[8]。
應對光照差異
[編輯]植物的葉具有極強的可塑性,它們的生長可能受光照水平的影響。在光照條件下生長的葉子往往更厚,這使得直射光下的光合作用效用最大化;葉子的面積更小,邊界層更薄,這使得葉子冷卻得更快。相反,生長在陰涼處的葉子往往更薄,有更大的表面積來捕獲更多的有限光線。[9][10]
應對土壤差異
[編輯]還有研究發現,在擬南芥、小鹽芥等植物中,根系中的運輸蛋白也隨著土壤鹽分和養分濃度的變化而發生改變[11]。如冰葉日中花等在內的一些植物在受到水分或鹽分短缺影響時,能夠改變它們的光合作用途徑,從而減少用水量[12]。
預測植物性狀
[編輯]由於表型可塑性的存在,在自然條件下生長的植物的性狀很難解釋和預測,除非能夠獲得一個明確的環境指數來量化環境:利用關鍵生長期的光熱時間鑑定高粱開花時間即屬於這樣的量化預測[6]。
動物中的例子
[編輯]現有研究已經顯示出營養和溫度對動物發育的影響[13],此外動物的行為也會因為食物的所在地[14]、母親在幼體幼年的撫摸行為[15]等多方面因素而受到影響[16]。由於物種的可塑性,動物能夠根據氣候和環境表現出不同的生存策略和覓食行為[17]。

應對溫度變化
[編輯]熱適應引起表型調整在不同類群中很常見。變溫動物需要對溫度變化進行自我調節,因為其生理的各個方面都直接取決於它們的熱環境。溫度變化可以通過影響甘油磷脂的脂肪鏈的移動,影響細胞膜的流動性。維持膜的流動性對細胞功能至關重要,變溫動物通過調節細胞膜的磷脂組成,以改變膜內范德瓦爾斯力的強度,從而維持不同溫度下膜的流動性。[18]
溫度變化還會對性別決定發生影響,許多爬行類的性別決定與溫度有關[19]:龜鱉類存在兩種溫度依賴的性別決定模式,受控於一定的溫度敏感期,一種是即低溫產雄性、高溫產雌性,另一種則為中間溫度產雄性而此外產雄性。DMRT1基因是首個被發現與這類性別決定有關係的基因,使用紅耳龜和太平洋麗龜作實驗,發現該基因的表達是溫度依賴的:在產雄性溫度和產雌性溫度下孵化,在胚胎的生殖嵴和中腎複合物均發現該基因的表達;溫度敏感期的後期則發現產雄性溫度發育胚胎性腺的表達水平較高,進一步的原理則有待研究[20]。隨著全球氣候變暖,生態學者已經在澳大利亞等地觀測到了雄性海龜孵化數量的下降[21]。
應對飲食變化
[編輯]消化系統的表型可塑性可以讓一些動物對飲食營養成分[22][23]、飲食質量[24][25]、能量需求[26][27][28]的變化做出反應。
脂類、蛋白質和碳水化合物的比例在內的飲食營養成分,可能會在如斷奶等的發育過程中發生變化,或隨著不同食物類別的豐富程度的季節性變化而發生變化。這些飲食變化可以引起小腸毛刷邊緣特定消化酶活性的可塑性。例如,在孵化後的最初幾天,家麻雀從高蛋白和高脂肪的昆蟲食物過渡到主要含有碳水化合物的種子食物,期間其麥芽糖酶活性增加了兩倍[29]。動物被人為要求適應高蛋白食物期間,其用以消化蛋白質的氨肽酶N的活性會增加。[30][31]
應對寄生蟲
[編輯]感染寄生蟲可以引發表型差異,以補償由寄生蟲引起的有害影響。通常無脊椎動物對寄生蟲去勢或毒力增加的反應是生殖力補償,例如:水蚤(學名:Daphnia magna)在接觸微孢子蟲的早期階段產生更多的後代,以補償未來生殖力下降帶來的適應度損失[32]。繁殖力的下降也可能是將營養物質重新導向以維持免疫反應[33]或減少其危害[34]的一種手段。在某些情況下,這種特殊形式的可塑性是由宿主派生的分子介導的[35]。
除繁殖外,宿主還可以通過生理上的可塑性來應對寄生。感染腸道線蟲的小鼠體驗到降低了葡萄糖在腸道的轉運率。為了彌補這一缺陷,小鼠增加了腸道黏膜細胞的總量,這些細胞負責葡萄糖的轉運。這使得受感染的小鼠保持與未受感染的小鼠相同的葡萄糖攝取能力和體型。[36]
表型可塑性也可以通過行為的變化來觀察。脊椎動物和無脊椎動物會傾向於自我治療感染,這可以被認為是一種適應性可塑性。[37]感染腸道蠕蟲的各種非人靈長類動物會吞食整片粗糙的葉子,從而將腸道中的寄生蟲排出體外。此外,葉刺激胃黏膜,促進胃酸分泌,增加腸道蠕動,可以促進消化道有效沖洗寄生蟲。[38]
應對捕食者
[編輯]
紅眼樹蛙(學名:Agalychnis callidryas)是一種棲息在中美洲熱帶地區的樹蛙。與許多青蛙不同的是,紅眼樹蛙有樹棲的卵,這些卵被產在池塘或水坑邊的樹葉上,懸掛於池塘或水坑上,孵化出來後,蝌蚪會掉進下面的水裡。這些樹棲卵遇到的最常見的食肉動物之一是是一種名為貓眼蛇(學名:Leptodeira septentrionalis)。當面臨捕食的直接威脅時,紅眼樹蛙能夠在產卵後5天提前孵化並在卵外存活:懸掛卵的連接結構感知周圍的震動中獲取信息,並判斷是否有被捕食的危險;在蛇攻擊的情況下,這一結構可以幾乎在一瞬間激活卵的孵化。已有的對照實驗顯示:當一個連接結構在六天大的時候受到攻擊時,整個連接結構所拖掛的卵幾乎是瞬間孵化出來的;而非危險狀態下,產卵後7天左右卵開始孵化到10天左右所有卵孵化結束;後續研究探索了紅眼樹蛙這一與孵化相關的表型可塑性的帶來的優勢和代價[39]。
表型可塑性和進化
[編輯]可塑性通常被認為是一種對環境變化的進化適應,它允許個體「較為適應」不同的環境,因而這種適應是可以合理預測的,並且發生在個體有機體的生命周期內。[40][41]如果一個特定環境中的最佳表型隨著環境條件的改變而改變,那麼個體表達不同特徵的能力應該是有利的,因此這一表型被自然選擇保留。因此,如果達爾文所說的適應度在改變表型後增加,這樣的可塑性在進化中逐漸占據多數。[42][43]
然而,可塑性對適應性的好處可能受到維持可塑性反應的能量成本制約,例如合成新的蛋白質、調整同工酶變異體的表達比例、維持感官機制以檢測變化都需要能量成本。同時,環境因素的的可預測性和可靠性亦限制了表型可塑性(見有利的適應性假設)。[44][45]
選擇實驗和進化實驗等方法的結果表明,可塑性既可以是一種在直接選擇條件(如捕食)下可以進化的性狀,也可以是對特定性狀平均值(如溫度變遷)的選擇的相關反應。[46]
適應性與適應不良
[編輯]應對環境產生的表型可塑性既可以是適應環境的,也可以在某種情況下轉變為不適應環境的。例如,淡水蝸牛(學名:Physa virgata)的蝸牛殼生長的可塑性既表現出了適應性,也表現出適應不良。在捕食者藍鰓太陽魚的存在下,這些蝸牛使它們的殼變得更圓,從而減少了生長,這使得它們具有更強的抗壓能力,更好地防止被捕食。然而,這些蝸牛並不能區分食肉太陽魚和非食肉太陽魚之間的化學信號差異,因而蝸牛對非食肉太陽魚作出改變了殼的形狀、減少生長的反應是不適當的,在食肉太陽魚缺失的情況下,這些變化限制了它們的繁殖力,且使蝸牛容易受到其他捕食者的影響。因此,這些淡水蝸牛對環境的適應性或不適應性反應取決於食肉太陽魚是否真的存在。[47][48]
氣候變異假說
[編輯]由於溫度具有深遠重要的生態影響,且在大範圍空間和時間尺度上溫度之變遷是可預測的,因此對熱量差異產生的適應已被認為是決定生物體表型可塑能力的一個關鍵機制。[49]溫度變化的幅度被認為與塑性能力成正比,因此在恆定的溫暖熱帶氣候中進化的物種,其塑性能力低於那些生活在天氣多變的溫帶氣候中的物種,這一假說被稱為「氣候變異假說[50](英語:Climatic variability hypothesis,简称CVH)」,並且已經獲得了一些關於植物和動物在整個緯度上的可塑性能力的研究的支持。[50][51][52]然而,對果蠅物種的研究未能發現一個明確的超過緯度梯度的可塑性模式,這表明這一假說可能不適用於所有分類族群或所有特徵。[53]一些研究人員提出,使用降水等因素直接測量環境變化,比單獨測量緯度更能預測表型可塑性。[54]
表型可塑性和氣候變化
[編輯]由於人類活動,預計今後100年內,氣候變化的速度將達到前所未有的程度。[55]表型可塑性允許生物個體在其一生中對氣候變化做出反應,因而成為生物應對氣候變化的關鍵機制。[56] 這一可塑性對於世代持續時間較長的物種尤為重要,因為通過自然選擇產生的進化可能不夠快,難以減輕氣候變暖的影響。
北美紅松鼠的繁殖期
[編輯]在過去的十年裡,北美紅松鼠(學名:Tamiasciurus hudsonicus)經歷了近2攝氏度的平均氣溫上升。溫度的上升導致了白雲杉球果數量的增加,而白雲杉球果是冬春季節繁殖的主要食物來源,食物的豐富導致北美紅松鼠的平均分娩日期提前了18天。食物豐富度對雌性個體的繁殖期有顯著影響,表明出表型可塑性。[57]
參見
[編輯]參考文獻
[編輯]- ^ The role of phenotypic plasticity in driving genetic evolution. Proceedings: Biological Sciences. July 2003, 270 (1523): 1433–40. PMC 1691402
 . PMID 12965006. doi:10.1098/rspb.2003.2372.
. PMID 12965006. doi:10.1098/rspb.2003.2372.
- ^ Kelly, Scott A.; Panhuis, Tami M.; Stoehr, Andrew M. Comprehensive Physiology. 2012: 1417–39. ISBN 978-0-470-65071-4. PMID 23798305. doi:10.1002/cphy.c110008.
- ^ 薛憲詞. 於黎. 昆虫非遗传多型性研究进展. 遺傳. 2017年9月, 39 (9): 798-809 [2019-04-28]. doi:10.16288/j.yczz.17-009. (原始內容存檔於2020-04-10).
- ^ Schlichting, CD. The Evolution of Phenotypic Plasticity in Plants. Annual Review of Ecology and Systematics. 1986, 17: 667–93. doi:10.1146/annurev.es.17.110186.003315.
- ^ International Aphid Genomics Consortium. Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biology. February 2010, 8 (2): e1000313. PMC 2826372 . PMID 20186266. doi:10.1371/journal.pbio.1000313.
- ^ 6.0 6.1 Li, Xin; Guo, Tingting; Mu, Qi; Li, Xianran; Yu, Jianming. Genomic and environmental determinants and their interplay underlying phenotypic plasticity. Proceedings of the National Academy of Sciences. 2018-06-07, 115 (26): 6679–6684. ISSN 0027-8424. PMC 6042117 . PMID 29891664. doi:10.1073/pnas.1718326115 (英語).
- ^ Silvertown, Jonathan. The paradox of seed size and adaptation. Trends in Ecology & Evolution. 1989, 4 (1): 24–26. PMID 21227308. doi:10.1016/0169-5347(89)90013-x.
- ^ Phenotypic plasticity for plant development, function and life history. Trends in Plant Science. December 2000, 5 (12): 537–42. PMID 11120476. doi:10.1016/S1360-1385(00)01797-0.
- ^ Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Functional Ecology. 2006, 20 (2): 207–16. JSTOR 3806552. doi:10.1111/j.1365-2435.2006.01105.x.
- ^ Lambers, Hans; Poorter, Hendrik. Advances in Ecological Research Volume 23. Advances in Ecological Research 23. 1992: 187–261. ISBN 978-0-12-013923-1. doi:10.1016/S0065-2504(08)60148-8.
- ^ Alemán, Fernando; Nieves-Cordones, Manuel; Martínez, Vicente; Rubio, Francisco. Differential regulation of the HAK5 genes encoding the high-affinity K+ transporters of Thellungiella halophila and Arabidopsis thaliana. Environmental and Experimental Botany. 2009, 65 (2–3): 263–9 [2019-04-30]. doi:10.1016/j.envexpbot.2008.09.011. (原始內容存檔於2020-07-24).
- ^ Tallman, Gary; Zhu, Jianxin; Mawson, Bruce T.; Amodeo, Gabriella; Nouhi, Zepedeh; Levy, Kathleen; Zeiger, Eduardo. Induction of CAM in Mesembryanthemum crystallinum Abolishes the Stomatal Response to Blue Light and Light-Dependent Zeaxanthin Formation in Guard Cell Chloroplasts. Plant and Cell Physiology. 1997, 38 (3): 236–42. doi:10.1093/oxfordjournals.pcp.a029158.
- ^ Weaver ME, Ingram DL. Morphological Changes in Swine Associated with Environmental Temperature. Ecology. 1969, 50 (4): 710–3. JSTOR 1936264. doi:10.2307/1936264.
- ^ Tremblay, Yann. Geographic variation in the foraging behaviour, diet and chick growth of rockhopper penguins (PDF). Marine Ecology. 2003. (原始內容 (PDF)存檔於2017-08-09).
- ^ Meaney, Michael J.; Szyf, Moshe; Dymov, Sergiy; Seckl, Jonathan R.; Sharma, Shakti; D'Alessio, Ana C.; Champagne, Frances A.; Cervoni, Nadia; Weaver, Ian C. G. Epigenetic programming by maternal behavior. Nature Neuroscience. 2004-08, 7 (8): 847–854 [2019-04-30]. ISSN 1546-1726. doi:10.1038/nn1276. (原始內容存檔於2021-05-07) (英語).
- ^ Web Topic 25.3 Environmental Induction of Behavioral Phenotypes. Companion Website for Developmental Biology, Eleventh Edition. [2019-04-30]. (原始內容存檔於2019-05-12).
- ^ Tremblay, Yann. Geographic variation in the foraging behaviour, diet and chick growth of rockhopper penguins (PDF). Marine Ecology. 2003. (原始內容 (PDF)存檔於2017-08-09).
- ^ Thermal adaptation in biological membranes: is homeoviscous adaptation the explanation?. Annual Review of Physiology. 1995, 57: 19–42. PMID 7778864. doi:10.1146/annurev.ph.57.030195.000315.
- ^ R. Shine; Warner, D. A. The adaptive significance of temperature-dependent sex determination in a reptile. Nature. 2008-01, 451 (7178): 566–568 [2019-04-30]. ISSN 1476-4687. doi:10.1038/nature06519. (原始內容存檔於2021-04-03) (英語).
- ^ 賀斌;史海濤;廖廣橋. 龟鳖类温度依赖型性别决定机制的研究进展. 動物學雜誌. 2009, 44 (5): 147~152 [2019-04-30]. (原始內容存檔於2022-05-11).
- ^ Craig Welch (撰文); 邱淑慧 (編譯). 為什麼這些海龜有99%轉變為雌性?. 國家地理中文網. 2018-01-30 [2019-04-30]. (原始內容存檔於2021-04-03).
- ^ Brzek P, Kohl K, Caviedes-Vidal E, Karasov WH. Developmental adjustments of house sparrow (Passer domesticus) nestlings to diet composition. The Journal of Experimental Biology. May 2009, 212 (Pt 9): 1284–93. PMID 19376949. doi:10.1242/jeb.023911.
- ^ Cortés PA, Franco M, Sabat P, Quijano SA, Nespolo RF. Bioenergetics and intestinal phenotypic flexibility in the microbiotherid marsupial (Dromiciops gliroides) from the temperate forest in South America. Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology. October 2011, 160 (2): 117–24. PMID 21627996. doi:10.1016/j.cbpa.2011.05.014.
- ^ Starck JM. Phenotypic flexibility of the avian gizzard: rapid, reversible and repeated changes of organ size in response to changes in dietary fibre content. The Journal of Experimental Biology. November 1999, 202 (22): 3171–9 [2019-04-30]. PMID 10539966. (原始內容存檔於2019-09-13).
- ^ Liu QS, Wang DH. Effects of diet quality on phenotypic flexibility of organ size and digestive function in Mongolian gerbils (Meriones unguiculatus). Journal of Comparative Physiology B. July 2007, 177 (5): 509–18. PMID 17333208. doi:10.1007/s00360-007-0149-4.
- ^ Naya DE, Ebensperger LA, Sabat P, Bozinovic F. Digestive and metabolic flexibility allows female degus to cope with lactation costs. Physiological and Biochemical Zoology. 2008, 81 (2): 186–94. PMID 18190284. doi:10.1086/527453.
- ^ Krockenberger AK, Hume ID. A flexible digestive strategy accommodates the nutritional demands of reproduction in a free-living folivore, the Koala (Phascolarctos cinereus). Functional Ecology. 2007, 21 (4): 748–756. doi:10.1111/j.1365-2435.2007.01279.x.
- ^ Hammond KA, Wunder BA. The Role of Diet Quality and Energy Need in the Nutritional Ecology of a Small Herbivore, Microtus ochrogaster. Physiological Zoology. 1991, 64 (2): 541–67. JSTOR 30158190. doi:10.1086/physzool.64.2.30158190.
- ^ Brzek P, Kohl K, Caviedes-Vidal E, Karasov WH. Developmental adjustments of house sparrow (Passer domesticus) nestlings to diet composition. The Journal of Experimental Biology. May 2009, 212 (Pt 9): 1284–93. PMID 19376949. doi:10.1242/jeb.023911.
- ^ Cortés PA, Franco M, Sabat P, Quijano SA, Nespolo RF. Bioenergetics and intestinal phenotypic flexibility in the microbiotherid marsupial (Dromiciops gliroides) from the temperate forest in South America. Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology. October 2011, 160 (2): 117–24. PMID 21627996. doi:10.1016/j.cbpa.2011.05.014.
- ^ Sabat P, Riveros JM, López-Pinto C. Phenotypic flexibility in the intestinal enzymes of the African clawed frog Xenopus laevis. Comparative Biochemistry and Physiology. Part A, Molecular & Integrative Physiology. January 2005, 140 (1): 135–9. PMID 15664322. doi:10.1016/j.cbpb.2004.11.010.
- ^ Chadwick W, Little TJ. A parasite-mediated life-history shift in Daphnia magna. Proceedings: Biological Sciences. March 2005, 272 (1562): 505–9. PMC 1578704 . PMID 15799946. doi:10.1098/rspb.2004.2959.
- ^ Ahmed AM, Baggott SL, Maingon R, Hurd H. The costs of mounting an immune response are reflected in the reproductive fitness of the mosquito Anopheles gambiae. Oikos. 2002, 97 (3): 371–377. doi:10.1034/j.1600-0706.2002.970307.x.
- ^ Hurd H. Host fecundity reduction: a strategy for damage limitation?. Trends in Parasitology. August 2001, 17 (8): 363–8. PMID 11685895. doi:10.1016/S1471-4922(01)01927-4.
- ^ Schallig HD, Hordijk PL, Oosthoek PW, Jong-Brink M. Schistosomin, a peptide present in the haemolymph of Lymnaea stagnal is infected with Trichobilharzia ocellata, is produced only in the snail's central nervous system. Parasitology Research. 1991, 77 (2): 152–6. doi:10.1007/BF00935429.
- ^ Kristan DM, Hammond KA. Physiological and morphological responses to simultaneous cold exposure and parasite infection by wild-derived house mice. Functional Ecology. 2003, 17 (4): 464–471. JSTOR 3598983. doi:10.1046/j.1365-2435.2003.00751.x.
- ^ Singer MS, Mace KC, Bernays EA. May RC , 編. Self-medication as adaptive plasticity: increased ingestion of plant toxins by parasitized caterpillars. PLOS One. 2009, 4 (3): e4796. PMC 2652102 . PMID 19274098. doi:10.1371/journal.pone.0004796.
- ^ Huffman MA. Self-Medicative Behavior in the African Great Apes: An Evolutionary Perspective into the Origins of Human Traditional Medicine. BioScience. 2001, 51 (8): 651–61. doi:10.1641/0006-3568(2001)051[0651:SMBITA]2.0.CO;2.
- ^ Adaptive plasticity in hatching age: a response to predation risk trade-offs. Proceedings of the National Academy of Sciences of the United States of America. April 1995, 92 (8): 3507–10. PMC 42196 . PMID 11607529. doi:10.1073/pnas.92.8.3507.
- ^ How stress selects for reversible phenotypic plasticity. Journal of Evolutionary Biology. 2005, 18 (4): 873–883. PMID 16033559. doi:10.1111/j.1420-9101.2005.00959.x.
- ^ Garland T, Kelly SA. Phenotypic plasticity and experimental evolution. Journal of Experimental Biology. 2006, 209 (12): 2344–2361. PMID 16731811. doi:10.1242/jeb.02244.
- ^ The genetics of phenotypic plasticity. V. Evolution of reaction norm shape. Journal of Evolutionary Biology. 1993, 6: 31–48. doi:10.1046/j.1420-9101.1993.6010031.x.
- ^ Evolution of phenotypic plasticity: patterns of plasticity and the emergence of ecotypes. The New Phytologist. April 2005, 166 (1): 101–17. PMID 15760355. doi:10.1111/j.1469-8137.2005.01322.x.
- ^ 周浩然. 徐明;於秀波;劉本彩. 生物适应性和表型可塑性理论研究进展. 西北農林科技大學學報(自然科學版). 2014年, 42 (4期): 215-220,228 [2019-04-28]. doi:10.13207/j.cnki.jnwafu.2014.04.006. (原始內容存檔於2020-04-10).
- ^ Dewitt, Thomas J; Sih, Andrew; Wilson, David Sloan. Costs and limits of phenotypic plasticity. Trends in Ecology & Evolution. 1998, 13 (2): 77–81. doi:10.1016/s0169-5347(97)01274-3.
- ^ Within-species digestive tract flexibility in rufous-collared sparrows and the climatic variability hypothesis. Physiological and Biochemical Zoology. 2011, 84 (4): 377–84. PMID 21743251. doi:10.1086/660970.
- ^ Langerhans, R. Brian; DeWit, Thomas J. Plasticity constrained: Over-generalized induction cues cause maladaptive phenotypes. Evolutionary Ecology Research. 2002, 4 (6): 857–70.
- ^ Costs and limits of phenotypic plasticity. Trends in Ecology & Evolution. February 1998, 13 (2): 77–81. PMID 21238209. doi:10.1016/S0169-5347(97)01274-3.
- ^ Janzen, Daniel H. Why Mountain Passes are Higher in the Tropics. The American Naturalist. 1967, 101 (919): 233–49. doi:10.1086/282487.
- ^ 50.0 50.1 李钧敏教授验证了中国入侵植物加拿大一枝黄花温度可塑性符合CVH假说,同时首次发现水分可塑性与经度密切相关. 浙江省植物進化生態學與保護重點實驗室. 2017-02-21 [2019-04-29]. (原始內容存檔於2019-05-18).
- ^ Latitudinal trends in digestive flexibility: testing the climatic variability hypothesis with data on the intestinal length of rodents. The American Naturalist. October 2008, 172 (4): E122–34. JSTOR 590957. PMID 18717635. doi:10.1086/590957.
- ^ Latitudinal patterns in phenotypic plasticity and fitness-related traits: assessing the climatic variability hypothesis (CVH) with an invasive plant species. PLOS One. 2012, 7 (10): e47620. PMC 3478289 . PMID 23110083. doi:10.1371/journal.pone.0047620.
- ^ Thermal tolerance in widespread and tropical Drosophila species: does phenotypic plasticity increase with latitude?. The American Naturalist. October 2011,. 178 Suppl 1: S80–96. PMID 21956094. doi:10.1086/661780.
- ^ Clements, Forrest E. The End of a World by Claude Anet. American Anthropologist. 1928, 30 (1): 125. JSTOR 660970. doi:10.1525/aa.1928.30.1.02a00120.
- ^ IPCC, 2014:氣候變化2014:綜合報告。政府間氣候變化專門委員會第五次評估報告第一工作組、第二工作組和第三工作組報告 (頁面存檔備份,存於網際網路檔案館) [核心撰写小组、 R.K. Pachauri 和 L.A. Meyer (eds.)]。瑞士日內瓦 IPCC,共151頁。
- ^ Towards an integrated framework for assessing the vulnerability of species to climate change. PLoS Biology. December 2008, 6 (12): 2621–6. PMC 2605927 . PMID 19108608. doi:10.1371/journal.pbio.0060325.
- ^ Genetic and plastic responses of a northern mammal to climate change. Proceedings: Biological Sciences. March 2003, 270 (1515): 591–6. JSTOR 3558706. PMC 1691280 . PMID 12769458. doi:10.1098/rspb.2002.2224.
拓展閱讀
[編輯]- Developmental Plasticity and Evolution. Oxford University Press. 2003. ISBN 978-0-19-512234-3.
- The Flexible Phenotype: A Body-Centred Integration of Ecology, Physiology, and Behaviour. Oxford University Press. 2011. ISBN 978-0-19-164015-5. 參見: Garland, Theodore. The Flexible Phenotype: A Body-Centred Integration of Ecology, Physiology, and Behaviour. Animal Behaviour. 2011, 82 (3): 609–10. doi:10.1016/j.anbehav.2011.06.012.
外部連結
[編輯]- Special issue of the Journal of Experimental Biology concerning phenotypic plasticity (頁面存檔備份,存於網際網路檔案館)
- Developmental Plasticity and Evolution - review of the book (頁面存檔備份,存於網際網路檔案館) from American Scientist
- Isidro A. T. Savillo's Impermanence of Sexual Phenotypes from Biologybrowser (Thomson Reuters)
- Phenotypic Plasticity lecture from the Institute for the Development of Educational Applications
- Environmental Induction of Behavioral Phenotypes from Developmental Biology, Eleventh Edition (頁面存檔備份,存於網際網路檔案館)
